Learning about the evolution of plant families
The Family Beds restored
Order beds or Family beds have been part of Botanic Gardens almost since their inception. They provide a means to display the systematic arrangement and relationships of plants to one another. Before an understanding of evolution, this was either based upon a folk-taxonomy of growth forms (trees, shrubs, and herbs for example) or more practical divisions, such as medicinal usages or geographical origins.
Attempts to lay out the beds on more Natural arrangements reached their high point in Bentham and Hooker’s landmark publication, Genera plantarum (1862-1883). The preparation of this work pre-dated the publication of Charles Darwin’s Origin of Species, so this is essentially a non-evolutionary scheme. However the arrangement of families was based on characters which provided a sequence from what they took as the most primitive (the Buttercup family) to the most derived families (the Grass family) – the Scalae Naturae.
Like Linnaeus’ sexual system, Bentham & Hooker’s classification was based on a few easily seen characters, such as whether the petals are fused to one another forming a tubular flower, or can be pulled off the flower one at a time. Such a system is therefore easily learnt, and looking at just a few characters will place an unknown specimen into a particular category. This arrangement is still the best way to learn the plant families even if it is not an accurate representation of how evolution of flowering plants has happened.
In the new Family bed layout we have tried to retain the Bentham and Hooker where it is a useful means to learning the families. But at the same time we have made some changes that demonstrate the modern appreciation of the evolution of the plant families that we have learnt from studies of biochemistry, and in recent years from DNA analysis.
Understanding plant families is not a simple matter – some botanists spend their entire lives discussing and debating the subject – so don’t feel confused or overwhelmed. Below we give four levels of explanation, they gradually become more jargon-filled and detailed . . . Give them a go and stop when you feel you have learnt enough!
At the bottom of this page are a number of other useful resources for understanding flowering plant evolution, including an interactive version.





1. A quick overview of flowering plant evolution
‘There are just two basic patterns of flowers, those with spiral petals and those with a single ring, or whorl of petals SPIRALS AND WHORLS
There are just two basic patterns of flowers: those with spirals of petals we now know are the most primitive, while those with whorls of petals are derived from these spiral-flowered ancestors.
We have laid out our family beds to reflect this pattern: Spiralled flowers are at the centre near the sundial, while the whorled flowers radiate out in four great evolutionary lineages.
MEET THE ANCESTORS
We now know that some of the earliest flowering plant types are still alive and many are well known to us, such as Magnolias and Waterlilies.
One of their characteristics is having flowers with spirally arranged parts, or very reduced flowers like Aristolochia or Peppers.
Others like the peppers have strong tasting compounds to stop them being eaten!
‘All our flower parts are in threes, sixes or nines, but never fives !’
GANG OF THREE
The Lilies, Grasses, Palms and Orchids all have one amazing feature in common – they have their flower parts in multiples of 3.
The six petals of the tulip, lily or Iris flower are common to all these species.
Even the much reduced grass flower still has three anthers.
‘All our flower parts are in fives or fours, but never in threes !’
GANG OF FIVES
Nearly all other flowers have five petals (a few have 4, or 6), and these divide into two major groups:
Those in which each petal is separate, as in a wild rose flower, and
Those in which the petals are fused into a tube like a Primrose or a Foxglove.
The Glasnevin Order Beds, a Summary
THE ANCIENT ONES: Paleotrees and Paleoherbs
We used to think of the evolution of flowering plants as a linear progression. Today we see it as a radiation, with 4 major evolutionary pathways. Two of these lineages are relics, but still around today; the magnolias and laurels which make up the Paleo-trees (literally Ancient trees), while water lilies and the dutchman’s pipe family are classified in the Paleo-herbs (literally Ancient herbs).
FROM SPIRALS TO WHORLS:
Paleotrees and Paleoherbs have spirals of flower parts, poor division between sepals and petals, and even a gradation from petals to stamens in flowers such as the waterlilies. All the remaining radiating lineages have whorls of petals and sepals, usually in threes or fives.
- PALEOTREES: Some of the earliest fossils of flowering plants are virtually indisitinguishable from modern Magnolia flowers. It is a remarkable fact that the pollinators of Magnolia flowers today (Beetles) were the only flying insect group at the time (bees and butterflies evolved much later), so it seems that the flowers of these trees have retained the same structure, and the same pollinators for over 100 million years !
1 MAGNOLIACEAE
2 WINTERACEAE
3 LAURACEAE
4 CALYCANTHACEAE
PALEOHERBS: The ancient position of the water lilies has long been overlooked. Other paleoherbs are characterised by their unusual flowers (Aristolochia) aromatic oils (Pepper family, Illicium) and flowers with many petals and sepals that intergrade with one another (Schisandra).
5 NYMPHACEAE
6 ARISTOLOCHIACEAE
7 SAURURACEAE
8 ILLICIACEAE and SCHISANDRACEAE
MONOCOTS: Paleotrees, Paleoherbs and Monocots all share the characteristic of pollen grains with a single pore or groove, through which the pollen tube germinates. The other defining characteristics of the Monocots are the floral parts in threes, and the single cotyledon of the seed.
EUDICOTS (Including the ROSIDS & ASTERIDS):
The remaining lineage is the true dicots (Eu- = true). All Eudicots have 3-porate pollen, while the Paleo groups and Monocots all share monocolpate pollen. One of the more unexpected discoveries was that the Sacred Lotus (Nelumbo) was closely related to the Proteas and the Plane tree, and that these three, seemingly unrelated families, are closely related, and diverge from the earliest Buttercup-like groupings.
There are two major lineages within the Eudicots; one includes the rose family and their relatives, the rosids (in red), the other comprises the daisy family and their relatives, the asterids (in yellow).
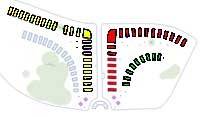
3. The New Synthesis – The Angiosperm Phylogeny Group
This diagram shows how the evolution of plants families as we understand them today matches the physical layout of the new Family beds at the National Botanic Gardens.
Angiosperm Phylogeny Group
On the far left are the MONOCOTS, which have all their flower parts in threes or multiples of threes. The first families, like the flowering rush, Butomus have many separate ovaries, as you progress along these beds, the ovaries of the flowers become fused into a single compound ovary, the anthers become reduced in number to 3 (Iris family) or 1 (Orchids and Gingers), and floral parts become reduced and minute (Grasses & Sedges).
Buttercups and Poppies are the most primitive of all the other evolutionary lines, unlike the Monocots, Paleotrees and Paleoherbs they have pollen grains with 3-pores as opposed to just a single pore. Plane trees and Proteas are closely related, and evolved from the buttercups at an early point in evolution. A number of other early offshoots of the Buttercup lineage are the CARYOPHYLLIDS (Pinks, Catchflies and Docks) and SAXIFRAGALES (Saxifrages, Sedums and Paeonieas).
The ROSIDS have separate petals that can be pulled off the flower one at a time. The initial beds, with the Cabbages and Violets have separate stamens also, while in families like the Mallows the anthers are fused into bundles. The Geraniums, Impatiens and Oxalis have their stamens fused to the base of the flower. The Peas, Roses and Passionflowers have the stamens fused to the Calyx. As you progress through these beds the number of flower parts reduces, and the ovaries move from being above the flower in Buttercups, to being completely hidden and below the flower in Fuchsias and the Carrot family.
The ASTERIDS have their petals fused together, making a tubular flower. The Daisy family and Campanulas have ovaries that lie below the flower, while in the Heather family onwards, the ovaries are contained inside the corolla tube. The Heather and Primrose families are characterised by having twice as many anthers as petals. From the corner bed onwards all the flowers have an ovary with just two compartments. Families like the mints at the very end of these beds have the fewest number of floral parts, with just 2 or 4 anthers per flower. These flowers are very specialised and efficient at being cross pollinated.
4. The Bentham and Hooker system (with APG modifications)
The Bentham and Hooker system is in blue, and where it differs substantially, the Angiosperm Phylogeny Group system is given in green.
Bentham and Hooker devised an essentially linear sequence of 200 families. To place a plant in this scheme, it is first necessary to decide which class it is; either a Dicotyledon or a Monocotyledon. This primary division of flowering plants was first proposed by John Ray (1628-1705). The Dicotyledons enlarge their stems by adding an outer layer (the growth rings of a tree = Exogenous growth), and the embryo has two cotyledons (like the two halves of a pea or broad bean); while in monocotyledons, new bundles are inserted throughout the stem cortex (Endogenous growth) and the embryo has just one cotyledon (The ‘germ’ of a wheat grain).
DICOTYLEDONS
Bentham & Hooker divided the Dicotyledons into three further divisions: two in which the flowers are furnished with both a calyx and a corolla (two perianth series), and in which the petals are separate (Poly-petalae), or fused into a tube (Gamo-petalae or Tubi-florae), and lastly those in which the flowers have only one perianth series (i.e. Daphne, right), usually with no corolla, but sometimes it is the calyx that is lacking (Mono-chlamydeae = “One-coated”). In evolutionary terms this last category comprised species with very reduced flowers – so much so that we can no longer physically ‘see’ where they belong in the system.
The study of biochemistry, and more latterly DNA studies, have revealed where these plant families truly belong in the evolutionary scheme, and thus in our order beds this category has completely disappeared.
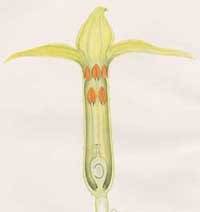
POLYPETALAE (=ROSIDS)
In turn the Polypetalae were divided into a further three sub-divisions: those where the petals, sepals and stamens are all separately attached to the receptacle (Thalami-florae, literally ‘bed flowers’); those with superior ovaries which are immersed in the disc of the flower (Disci-florae); and lastly those where the stamens are attached to the calyx of the flower (Calyci-florae).
THALAMIFLORAE: The Sepals, Petals and Stamens in these flowers are each separately attached to the receptacle.
Beds 21 to 28
21 RANUNCULACEAE
22 LARDIZABALACEAE
23 BERBERIDACEAE
23 PAPAVERACEAE
24 CRUCIFERAE
24 CAPPARACEAE, RESEDACEAE
25 CISTACEAE
25 VIOLACEAE, POLYGALACEAE
27 HYPERICACEAE
27 MALVACEAE
28 TILIACEAE
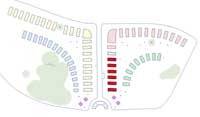
DISCIFLORAE:
In these flowers the ovary is superior, but is immersed, or sunken, into the disk of the flower.
Beds 29 to 34
29 LINACEAE
29 LIMNANTHACEAE
30 GERANIACEAE
31 BALSAMINACEAE
31 OXALIDACEAE
31 VITACEAE
32 RUTACEAE
33 AQUIFOLIACEAE
34 RHAMNACEAE
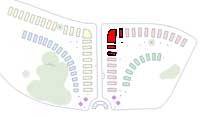
CALYCIFLORAE:
The Stamens are fused to the Calyx of the flower, often forming a hypanthium, a tube below the petals and sepals, but above the ovary, which may be superior or inferior.
Beds 35 to 48
35/36 LEGUMINOSAE
37/38 ROSACEAE
39 EUPHORBIACEAE
39 URTICACEAE
40 JUGLANDACEAE
41 BETULACEAE
42 SALICACEAE
43 MYRTACEAE
44 LYTHRACEAE
44 ONAGRACEAE
45 LOASACEAE
45 PASSIFLORACEAE
46 CUCURBITACEAE
46 BEGONIACEAE
47 UMBELLIFERAE
48 ARALIACEAE
Families like the Spurges (Euphorbiaceae), Walnuts (Juglandaceae), Birches (Betulaceae), Willows (Salicaceae), Stinging nettles (Urticaceae) and Elms have such reduced, wind-pollinated flowers, that Bentham and Hooker put them into their Monochlamydeae class. Today we know that they are in fact related to families like the roses (Rosaceae).
CARYOPHYLLIDS
In these families the ovary placentation is centrally positioned in the ovary, either being axile, or basal, and the seeds have a characteristic curved embryo. Because of their highly reduced flowers, which lack both sepals and petals, Bentham & Hooker classified many of these in their Monochlamydae.
Beds 11 to 15
11 CARYOPHYLLACEAE
12 PORTULACACEAE
12 AIZOACEAE
13 NYCTAGINACEAE
13 AMARANTHACEAE
14 CHENOPODIACEAE
14 PHYTOLACCACEAE
15 POLYGONACEAE
SAXIFRAGALES
These were formerly classified as being close to Rosaceae and Leguminosae, but are a distinct and early lineage of Eudicots. They have partly-fused ovaries.
Beds 16 to 20
16 SAXIFRAGACEAE
17 PAEONIACEAE
18 CRASSULACEAE
19 GROSSULARIACEAE
20 HAMAMELIDACEAE
GAMOPETALAE (=ASTERIDS) (MAIN TITLE)
The Gamopetalae (=fused petals) are divided into those with an inferior ovary (Inferae); those with a superior ovary, and in which the stamens are opposite the petals or double the number of petals (Hetero-merae = unequal numbers); and lastly those with superior ovaries with 2 carpels (Bi-carpellatae = two carpels).
49 CORNACEAE
INFERAE: The Ovary is below the flower (inferior), and the number of stamens is equal to the number of petals.
Beds 50 to 55
50 CAPRIFOLIACEAE
50 RUBIACEAE
51 VALERIANACEAE
52 DIPSACACEAE
53/54 COMPOSITAE
55 CAMPANULACEAE
HETEROMERAE : The Ovary is within the flower (superior) and the number of stamens is either double the number of petals, or each stamen is aligned on the centre-line of the petal.
Beds 56 to 60 56 ERICACEAE
57 PLUMBAGINACEAE
58 PRIMULACEAE
59 EBENACEAE
60 STYRACACEAE
BICARPELLATAE: The Ovary is within the flower (superior) and is divided into 2 carpels (these look like 4 in Boraginaceae and Lamiaceae).
Beds 61 to 74 61 OLEACEAE
62 APOCYNACEAE
62 ASCLEPIADACEAE
63 GENTIANACEAE
64 POLEMONIACEAE
64 HYDROPHYLLACEAE
65 BORAGINACEAE
66 CONVOLVULACEAE
67 SOLANACEAE
68/69 SCROPHULARIACEAE
70 ACANTHACEAE
71 GLOBULARIACEAE
71 BIGNONIACEAE
72 VERBENACEAE
73/74 LABIATAE
74 PLANTAGINACEAE
MONOCOTYLEDONS
The family Liliaceae used to be classified as a single large family (78-86), today it is divided into some 20 separate families, each distinguished by simple structural differences. Bentham & Hooker’s arrangement of families within this division is long outdated, and the new arrangment followed here relates to many morphological features of growth.
Beds 75 to 90 These families have separate carpels
75 ARACEAE
76 ALISMATACEAE and BUTOMACEAE
These are isolated and distinctive families
77 ORCHIDACEAE
77 IRIDACEAE
These families have essentially similar calyx & corolla lobes
78 HEMEROCALLIDACEAE and PHORMIACEAE
79 HYACINTHACEAE and THEMIDACEAE
80 AGAPANTHACEAE, ALLIACEAE and AMARYLLIDACEAE
81 HOSTACEAE and AGAVACEAE
82 ASPARAGACEAE and ASPHODELLACEAE
83 ASTELIACEAE, CONVALLARIACEAE and RUSCACEAE
84/85 LILIACEAE
85 SMILACACEAE
86 ALSTROEMERIACEAE, COLCHICACEAE and MELANTHIACEAE
These families have calyx and corolla lobes that are not alike
87 PALMAE and BROMELIACEAE
88 JUNCACEAE and CYPERACEAE
89 GRAMINEAE
90 COMMELINACEAE and ZINGIBERACEAE
overview of the beds
Paleotrees and Paleoherbs
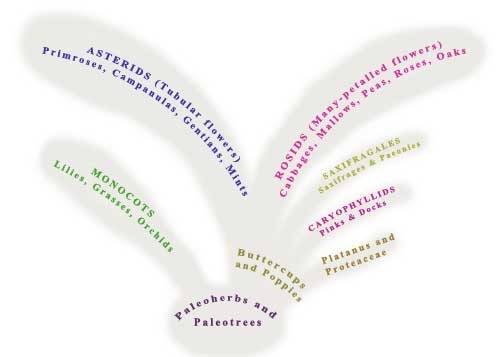
Magnolia (top left) Waterlily (top-right). If you move your mouse over the image, you will also see other members of this ancient grouping which are characterised by their spirally arranged petals.
On the lawn by the Sundial are a Magnolia and a member of the Laurel fmily. These trees evolved very early on in flowering plant evolution, and we refer to them ass belonging to the Paleo-trees (Ancient trees). On the surrounding lawns are four smaller beds, which represent the Paleo-herbs (Ancient herbs) these are non-woody plant families including waterlilies and the Pepper Family (Piperaceae).
Remarkably Magnolia flowers are still almost identical to those of trees growing 80 million years ago. Along with Waterlilies, the flowers of Magnolia are still pollinated by beetles, these were the only herbivorous flying insects that existed at the time, butterflies, moths, bees and wasps only evolved much later.

Rosids
Apart from the monocotyledons (see below) all the remaining flowering plants evolved from a Buttercup-like ancestor. Fowers from this group have separate petals that can be pulled off the flower one at a time. Unlike the Paleo-trees, paleo-herbs and monocts, all the species in these groupings have pollen grains with 3 pores and not a single pore.
The initial beds, with the Cabbages (Brassicaceae) and Violets (Violaceae) have separate stamens also, while in families like the Mallows (Malvaceae) the anthers are fused into bundles. The Geraniums (Geraniaceae), Impatiens (Baslsaminaceae) and Oxalis (Oxalidaceae) have their stamens fused to the base of the flower. The Peas, Roses and Passionflowers have the stamens fused to the Calyx. As you progress through these beds the number of flower parts reduces, and the ovaries move from being above the flower in Buttercups, to being completely hidden and below the flower in Fuchsias (Onagraceae) and the Carrot family (Apiaceae).

Asterids
The Asterids have their petals fused together, making a tubular flower. The Daisy family (Asteraceae), Campanulas (Campanulaceae) and Valerians (Valerianaceae) have ovaries that lie below the flower, while in the Heather family (Ericaceae) onwards, the ovaries are contained inside the corolla tube. The Heather and Primrose (Primulaceae) families are characterised by having twice as many anthers as petals. From the corner bed onwards all the flowers have an ovary with just two compartments. Families like the mints at the very end of these beds have the fewest number of floral parts, with just 2 or 4 anthers per flower. These flowers are very specialised and efficient at being cross pollinated.

Saxifragales and Caryophyllids
The pinks and catchflys (Caryophyllaceae) are closely related to the docks and knotweeds (Polygonaceae). The ovary either has a central axis on which the ovules are borne, or these are all basal. They divide into two main groups – those with curved embryos in their seeds (Amaranthaceae, Cactaceae, Caryophyllaceae and Chenopodiaceae) or those with straight embryos (Plumbaginaceae and Polygonaceae). Most are herbaceous and those that are woody have anomalous secondary growth.
The Saxifrages used to be classified close to the Rosaceae, while Paeonies were at one time placed with the Buttercups (Ranunculaceae). We now know they are better placed together, characterised by their half fused ovaries.

Monocotyledons

ANGIOSPERM PHYLOGENY
A pdf summary of Bentham and Hooker’s divisions of the plant families as recognised by them in the 1860s (Genera Plantarum). Essentially this is a pre-evolutionary, natural system, and in essence is what the naked eye can recognise. The Angiosperm Phylogeny system is summarised as an evolutionary tree of all the flowering plant families based upon DNA evidence.
The most interesting fact is how the morphological and molecular evidence supports one another.
PDF LINK IN DESKTOP FOLDER FOR THIS PAGE
A map and student’s guide to the Family beds.
[PDF LINK IN DESKTOP FOLDER FOR THIS PAGE]
